- Research
- Open access
- Published:
Global dynamics in a class of discrete-time epidemic models with disease courses
Advances in Difference Equations volume 2013, Article number: 57 (2013)
Abstract
In this paper, a class of discrete SIRS epidemic models with disease courses is studied. The basic reproduction number is computed. The main results on the permanence and extinction of the disease are established. That is, the disease-free equilibrium is globally attractive if , and there exists a unique endemic equilibrium and the disease is also permanent if .
MSC:39A30, 92D30.
1 Introduction
In recent years, more and more attention has been paid to the discrete-time epidemic models. There are several reasons for that. Firstly, since the statistic data about a disease is collected by day, week, month or year, it is more direct, more convenient and more accurate to describe the disease by using the discrete-time models than the continuous-time models; secondly, the discrete-time models have more wealthy dynamical behaviors; for example, the single-species discrete-time models have bifurcations, chaos and other more complex dynamical behaviors.
For a discrete-time epidemic model, we see that at the present time, the main research subjects are the computation of the basic reproduction number, the local and global stability of the disease-free equilibrium and endemic equilibrium, the extinction, persistence and permanence of the disease, and the bifurcations, chaos and more complex dynamical behaviors of the model, etc. Many important and interesting results can be found in articles [1–24] and the references cited therein.
In [4], the next generation matrix approach for calculating the basic reproduction number is summarized for discrete-time epidemic models. As applications, six disease models have been developed for the study of two emerging wildlife diseases: hantavirus in rodents and chytridiomycosis in amphibians. The comparison of deterministic and stochastic SIS and SIR type epidemic models in discrete time is discussed in [3]. In [8, 9], the discrete-time SIS type epidemic models with periodic environment and with disease-induced mortality in density-dependence, respectively, are investigated. In [11], Izzo and Vecchio proposed an implicit nonlinear system of difference equations which represents the discrete counterpart of a large class of continuous models concerning the dynamics of an infection in an organism or in a host population. They also studied the limiting behavior of the discrete model and derived the basic reproduction number. Izzo, Muroya and Vecchio in [10] proved the globally asymptotic stability of the disease-free equilibrium for a general discrete-time model of population dynamics in the presence of an infection. For the discrete epidemic model with immigration of infectives, by adopting the means of the nonstandard discretization method from continuous epidemic, Jang and Eiaydi in [12] studied the globally asymptotic stability of the disease-free equilibrium, the locally asymptotic stability of the endemic equilibrium and the strong persistence of the susceptible class. Li and Wang in [15] discussed a SIS type discrete epidemic model with stage structure, where Beverton-Holt type and Richer type recruitment rates were considered, the global stability of the disease-free equilibrium and the dynamical complexity were investigated. In [17], the sufficient and necessary conditions for the global stability of the endemic equilibrium were established for a discrete epidemic model for the disease with immunity and latency in a heterogeneous host population. In [19], the bifurcations and chaos were proved in a discrete epidemic model with nonlinear incidence rates. The permanence and extinction are investigated in [20–22] for a class of discrete SIRS and SEIRS type epidemic models with time delays. In [24], a discrete mathematical model is formulated to investigate the transmission and control of SARS in China, where the basic reproductive number is obtained as a threshold to determine the asymptotic behavior of the model. Particularly, in [18] the authors studied the following class of disease epidemic models with the spread of an infection in a host population:

The global stability of disease-free equilibrium and endemic equilibrium and the permanence of the disease were obtained.
However, we know that many diseases have different disease courses, for example, tuberculosis, syphilis, AIDS, etc. Therefore, taking into account the epidemic models with disease courses is very important since disease pathogen bacteria with different course may have different reproduction and survival capacities, which indirectly influences the population growth. Under a different disease course, the transmission rate, the mortality and other vital parameters will be different [25–27].
Motivated by the above results, in this paper, we consider a class of discrete-time epidemic models with disease courses. We divide the total population into subgroups according to m disease courses. Let be the number of susceptible individuals at the n th generation, () denote the number of infectious individuals who are in the j th course of a disease at n th generation, and let denote the number of recovered individuals at the n th generation. We introduce the following assumptions.
-
(1)
The susceptible x has a constant input rate Λ and a natural death rate d.
-
(2)
The susceptible individuals of the th generation are only infected by the infectious individuals of the n th generation, and is the constant transmission coefficient of which the susceptible is infected by compartment .
-
(3)
After a susceptible individual contacts infectives and is infected, he/she will firstly enter compartment , and then turn into compartments , finally into compartment .
-
(4)
The infectious in the j th disease course admits the constant natural death rate d, the constant death rate induced by disease , the constant recovery rate and the constant transmit rate from compartment to .
-
(5)
The recovered z admits the constant natural death rate d, does not have permanent immunity, hence there is a constant transfer rate δ from the recovered class back to the susceptible class.
Base on the above assumptions, a class of discrete-time epidemic dynamical models with m disease courses can be established as follows:
where , , . For model (1), we always assume that the following basic hypotheses hold.
(H1) For each , , , , , , , , and .
(H2) Any solution of model (1) satisfies the following initial conditions:
Remark 1 For model (1), we can easily see that when then model (1) describes a discrete SIRS type epidemic model with disease courses, where () denotes the number of infectious individuals in the i th course of the disease; and when then model (1) describes a discrete SEIRS type epidemic model with disease courses, where is exposed and () denotes the number of infectious individuals in the i th course of the disease.
Remark 2 In model (1), based on the above assumption (1), we know that the disease incidence term is denoted by . This makes , i.e., the susceptible number of the th generation, appear on both sides of the first equation. The reason for the above arguments is based on two considerations. On the one hand, it is influenced by the works given in [11, 18]; on the other hand, for the sake of convenience for mathematical analysis, especially, the positivity of solutions in model (1).
In this paper, by developing the methods given in [10, 11, 18], we will give the explicit expression of the basic reproduction number . The criteria on the permanence and extinction of the disease will be established. That is, the disease-free equilibrium is globally attractive if , and there exists a unique endemic equilibrium and the disease is also permanent if .
This paper is organized as follows. In Section 2, as preliminaries we will give several lemmas which will be used in the proofs of the main results. In Section 3, the basic reproduction number is calculated, the existence on the disease-free equilibrium and endemic equilibrium is given and the theorem on the globally asymptotic stability of the disease-free equilibrium is stated and proved. In Section 4, we will obtain the permanence of the disease. Conclusions are presented in the last section.
2 Preliminaries
Let k be any positive integer, we denote . For any sequence , we define
Firstly, on the positivity of solutions of model (1), we have the following result.
Lemma 1 For any solution of model (1), it holds that , , () for all .
Proof From model (1), we can easily obtain

and
Assume that , () and , then we further have

and
Therefore, by using the induction, we get , , for all and . This completes the proof. □
Lemma 2 For any solution of model (1), it follows that
Proof
Let
then we have
By using the induction, we can obtain the following inequality:
from which we have
From this, we finally have
This completes the proof. □
On the weak permanence and permanence of the disease of model (1), we have the following definitions.
The disease in model (1) is said to be weak permanent (permanent) if there exists a constant such that, for any solution sequence of model (1), one has
From Lemma 2, Theorem 1.1.3 and Theorem 1.10 given in [28], we can immediately obtain the following result.
Lemma 3 If the disease in model (1) is weak permanent, then it also is permanent.
Similar to Lemma 2.3 in [11] and Lemma 5 in [18], we have the following result.
Lemma 4 For any solution of model (1), the following inequalities hold:


Meanwhile, we also have
where
Proof From model (1) and Lemmas 1 and 2, we easily have that
and then
From the third equation of model (1), we directly have
Hence, we immediately obtain that for .
Considering the second equation of model (1), we can obtain the following inequality:
and
Then we have
Similarly, from model (1), we easily obtain
and
Hence, from (7) and (8), it can be easily proved that
Hence, inequality (5) holds.
From inequalities (7), (8) and (9), it follows that
and
Hence, inequality (6) holds. This completes the proof. □
3 Global attractivity of disease-free equilibrium
Let the constant
Firstly, on the existence of disease-free equilibrium and endemic equilibrium, we have the following result.
Theorem 1 (1) Model (1) always has a disease-free equilibrium .
-
(2)
When , model (1) also has a unique endemic equilibrium , where
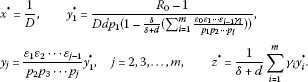
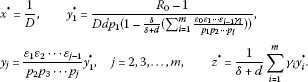
Proof The equilibrium of model (1) satisfies the following equations:
From the third equation to -equation of (11), we easily obtain
Substituting (12) into the second equation of (11), we further have
Having solved this equality, we obtain that and .
When , then from (12) we have for . Further, from the first and the last equations of (11), we have and . This shows that model (1) has a disease-free equilibrium .
When , then from the last equation of (11), we have
From the first equation of (11), we further obtain
Substituting (12) into this equality, we further have

Hence,
Then, we further have
Thus, we finally obtain
Since for each and , we have

Then we further obtain

Hence, we can infer that
Thus, from (14) we obtain that if and only if . Further, from (12) we obtain for . Finally, from (13), we also have . Therefore, we prove that model (1) has a unique endemic equilibrium . This completes the proof. □
Remark 3
Obviously, we have
The first term denotes the ultimate number of the susceptible at the end of the first disease course which is infected by an infectious individual of the first disease course. The second term denotes the ultimate number of the susceptible at the end of the second disease course which is infected by an infectious individual of the second disease course. And lastly, the final term denotes the ultimate number of the susceptible at the end of the m th disease course which is infected by an infectious individual of the m th disease course. We see that is the sum of these ultimate numbers. This shows that certainly is the basic reproduction number of model (1).
Theorem 2 If , then the disease-free equilibrium of model (1) is globally attractive. That is, for any solution of model (1), we have
Proof Since , then by inequality (6) in Lemma 4, we can obtain (). In fact, if for some such that , then by inequality (4) in Lemma 4, we can obtain . Hence, . From inequality (6) in Lemma 4 and , we have
which leads to a contradiction. Hence, (). Finally, from the expression of and of model (1), we can infer that (15) holds. This completes the proof. □
4 Permanence of disease
In this section, we mainly prove the permanence of model (1) when . Firstly, we introduce several lemmas which will be used to study the permanence of model (1). Consider the following auxiliary system:
where is a given non-negative bounded real sequence, and parameters , , , , δ and d (, ) are defined as in model (1). We have the following result.
Lemma 5 For any constants and , there exist a constant and an integer such that for any initial time and initial value with (), if for all , then we have
where is the solution of system (16) with the initial condition .
Proof Firstly, we consider the last equation of system (16)
We can obtain that for any constants and , there exist and , with
such that for any initial time and initial value , if for all and , then we have
Consider the m th equation of system (16)
For the above constants and , there exist a constant and an integer , with
such that for any initial time and initial value , if for all , then we have
Further consider the th equation
For the above constants and , there exist a constant and an integer , with
such that for any initial time and initial value , if for all , then we have
Repeating the above process for , respectively, finally we can obtain that for each () and for the above obtained constants and , there exist a constant and an integer , with
such that for any initial time and , if for all , then we have
Let . Then, from the above discussions, we obtain that for any initial time and initial value (), if for all , then from (18) we have
We further have
Lastly, from (17) we have
This shows
This completes the proof. □
We further consider the following equation:
where the parameters are assumed to be as in system (1) and . By calculating, we obtain that equation (19) has a positive equilibrium with
Obviously, we have
Hence, there exists an such that when , we have
Therefore, we have the following result.
Lemma 6 For any constants and , there exists an integer such that for any initial and initial value with , we have
where is the solution of equation (19) with the initial condition .
Proof For any solution of equation (19) with the initial condition , we define a function
then
From this, we easily see that for any constant ε, there exists an integer such that for any initial time and initial value with , when , then for all , we have
This shows that the conclusion of Lemma 6 holds. This completes the proof. □
For any with and , if (), then we denote .
Let and be a function defined on and . If for any with we have
then the function is said to be non-decreasing for .
Lemma 7 (See [29])
Let the domain and the function defined on and be non-decreasing for . If the sequence for all satisfies (), then we have
where is the solution of the difference equation with initial value ().
Now, we consider the following linear autonomous difference system:
where A is an non-negative matrix and . Then we have the following result.
Lemma 8 Let be the spectral radius of matrix A, then the following conclusions hold.
-
(1)
There exists an m-dimensional column vector with () such that is a solution of system (22).
-
(2)
For any with (), there exist constants () such that
where is the solution of system (22) satisfying the initial condition .
Proof (1) Since λ is an eigenvalue of matrix A and matrix A is a non-negative matrix, we can obtain that there is a vector corresponding to the eigenvalue λ such that
From , we have
Suppose that , then
Therefore, is a solution of system (22).
-
(2)
Let , , and . Further let and . Then we have . Let and . Then and are the solutions of system (22) with initial value and , respectively. Then from Lemma 7 it follows that
for all . □
Lemma 9 If , then there exists a constant such that for any solution of model (1), we have
Proof
Since
then we can choose a constant () such that
Hence, we have
Then, from (20) and (21), there exists an with such that
Now, from Lemma 6, for above , there exists an integer such that for any initial time and initial value with , where , we have
where is the solution of equation (19) with and the initial condition . Hence, from (25) and (26), we have
Assume that is any positive solution of model (1) with the initial condition . Then, from Lemma 2, for there exists an integer such that when , we have
Consider the following difference system:
where parameters , , , , δ and d in system (29) are given as in model (1). Since is the solution of model (1), then is the solution of system (29). From (28) and Lemma 5, for above and , there exist a constant and an integer with such that for any initial time and initial value , if for all , then we have for all and . Hence, if for all , then we have
Now, we prove that if then
for any positive solution of model (1). Suppose that the conclusion is not true, then there exists a positive solution of model (1) such that . Hence, there exists an integer such that for all . From (30) we know that for any , there exists an integer such that
Then, from (28) and the first equation of model (1), we obtain
Since from Lemma 7 and (27), for any , there exists an integer such that
then replacing (32) into the second and the third equations of model (1), we have
Next, consider the following auxiliary system:
Obviously, system (34) is a linear autonomous difference system and we can rewrite it as follows:
where and
We have
From (24), we have
and, for a constant , we further have
Then, from the intermediate value theorem, there exists a constant such that . Therefore, we can obtain . Let be the solution of system (33) with the initial condition , where . Then from (33), (34) and Lemma 7, we have
Further, from the second part of Lemma 8, there exists a constant such that
Hence, we have
From this, we obtain
which leads to a contradiction. This completes the proof. □
Lastly, directly from Lemma 3, Lemma 4 and Lemma 9, we can obtain the following result on the permanence of model (1).
Theorem 3 If , then the disease in model (1) is permanent.
Proof In fact, from Lemma 9, we obtain that for any positive solution of model (1),
Then, from inequality (4) in Lemma 4, we further have
Finally, from the last equation of model (1), we have
This completes the proof. □
5 Conclusions
In this paper, we study a class of discrete epidemic models with disease courses, that is, model (1). The basic reproduction number is calculated. It is shown that the global dynamics of model (1) is determined by the basic reproduction number . If , then we obtain that the disease-free equilibrium of model (1) is globally asymptotically stable. This shows that when the disease in model (1) is extinct. If , then we obtain that the endemic equilibrium of model (1) exists and the disease is permanent. Clearly, our condition given in this paper is the threshold condition between the extinction and the permanence of the disease. Hence, our results obtained in this paper extend the results given in [7, 11, 18] for the discrete epidemic models.
However, it is a pity that the case of the basic reproduction number is not discussed in this paper. From the results on the case obtained in [17, 18], we can guess that when then the disease-free equilibrium of model (1) is also globally asymptotically stable. This shows that when the disease in model (1) is also extinct. The other one which is not obtained in our this paper is the global stability of the endemic equilibrium of model (1). From the results on the global stability of the endemic equilibrium obtained in [7, 17, 18], we can guess that when the endemic equilibrium of model (1) is globally stable. We will discuss these problems in our future work.
References
Agarwal RP, O’Regan O, Wong PJY: Dynamics of epidemics in homogeneous/heterogeneous populations and the spreading of multiple inter-related infectious diseases: constant-sign periodic solutions for the discrete model. Nonlinear Anal., Real World Appl. 2007, 8: 1040-1061. 10.1016/j.nonrwa.2006.06.005
Allen LJS: Some discrete-time SI, SIR and SIS epidemic models. Math. Biosci. 1994, 124: 83-105. 10.1016/0025-5564(94)90025-6
Allen LJS, Burgin AM: Comparison of deterministic and stochastic SIS and SIR models in discrete time. Math. Biosci. 2000, 163: 1-33. 10.1016/S0025-5564(99)00047-4
Allen LJS, Driessche P: The basic reproduction number in some discrete-time epidemic models. J. Differ. Equ. Appl. 2008, 14: 1127-1147. 10.1080/10236190802332308
Allen LJS, Lou Y, Nevai AL: Spatial patterns in a discrete-time SIS patch model. J. Math. Biol. 2009, 58: 339-375. 10.1007/s00285-008-0194-y
Castillo-Chavez C, Yakubu AA: Discrete-time SIS models with complex dynamics. Nonlinear Anal. 2001, 47: 4753-4762. 10.1016/S0362-546X(01)00587-9
Enatsu Y, Nakata Y: Global stability for a class of discrete SIR epidemic models. Math. Biosci. Eng. 2010, 7: 347-361.
Franke LE, Yakubu AA: Discrete-time SIS epidemic model in a seasonal environment. SIAM J. Appl. Math. 2006, 66: 1563-1587. 10.1137/050638345
Franke LE, Yakubu AA: Disease-induced mortality in density-dependent discrete-time S-I-S epidemic models. J. Math. Biol. 2008, 57: 755-790. 10.1007/s00285-008-0188-9
Izzo G, Muroya Y, Vecchio A: A general discrete time model of population dynamics in the presence of an infection. Discrete Dyn. Nat. Soc. 2009., 2009: Article ID 143019. doi:10.1155/2009/143019
Izzo G, Vecchio A: A discrete time version for models of population dynamics in the presence of an infection. J. Comput. Appl. Math. 2007, 210: 210-221. 10.1016/j.cam.2006.10.065
Jang, S, Elaydi, SN: Difference equations from discretization of a continuous epidemic model with immigration of infectives. Technical report, V. 92, MTBI Cornell University (2004)
Li J, Lou J, Lou M: Some discrete SI and SIS epidemic models. Appl. Math. Mech. 2008, 29: 113-119.
Li J, Ma Z, Brauer F: Global analysis of discrete-time SI and SIS epidemic models. Math. Biosci. Eng. 2007, 4: 699-710.
Li X, Wang X: A discrete epidemic model with stage structure. Chaos Solitons Fractals 2005, 26: 947-958. 10.1016/j.chaos.2005.01.063
Oli MK, Venkataraman M, Klein PA, Wendland LD, Brown MB: Population dynamics of infectious diseases: a discrete time model. Ecol. Model. 2006, 198: 183-194. 10.1016/j.ecolmodel.2006.04.007
Muroya Y, Bellen A, Enatsu Y, Nakata Y: Global stability for a discrete epidemic model for disease with immunity and latency spreading in a heterogeneous host population. Nonlinear Anal., Real World Appl. 2012, 13: 258-274. 10.1016/j.nonrwa.2011.07.031
Muroya Y, Nakata Y, Izzo G, Vecchio A: Permanence and global stability of a class of discrete epidemic models. Nonlinear Anal., Real World Appl. 2011, 12: 2105-2117. 10.1016/j.nonrwa.2010.12.025
Papaschinopoulos G, Stefanidou G, Papadopoulos KB: On a modification of a discrete epidemic model. Comput. Math. Appl. 2010, 59: 3559-3569. 10.1016/j.camwa.2010.03.049
Sekiguchi M: Permanence for some discrete epidemic models. Int. J. Biomath. 2009, 2: 443-461. 10.1142/S1793524509000807
Sekiguchi M: Permanence of a discrete SIRS epidemic model with time delays. Appl. Math. Lett. 2010, 23: 1280-1285. 10.1016/j.aml.2010.06.013
Sekiguchi M, Ishiwata E: Global dynamics of a discretized SIRS epidemic model with time delay. J. Math. Anal. Appl. 2010, 371: 195-202. 10.1016/j.jmaa.2010.05.007
Zhang D, Shi B: Oscillation and global asymptotic stability in a discrete epidemic model. J. Math. Anal. Appl. 2003, 278: 194-202. 10.1016/S0022-247X(02)00717-5
Zhou Y, Ma M, Brauer F: A discrete epidemic model for SARS transmission and control in China. Math. Comput. Model. 2004, 40: 1491-1506. 10.1016/j.mcm.2005.01.007
Andersen BL, Kiecolt-Glaser LK, Glaser R: A biobehavioral model of cancer stress and disease course. Am. Psychol. 1994, 49(5):389-404.
Kastbom A, Strandberg G, Lindroos A, Skogh T: Anti-CCP antibody test predicts the disease course during 3 years in early rheumatoid arthritis (The Swedish TIRA project). Ann. Rheum. Dis. 2004, 63: 1085-1089. 10.1136/ard.2003.016808
Vingsbo C, Sahistrand P, Brun JG, Jonsson R, Saxne T, Holmdahl R: A new model for rheumatoid arthritis with a chronic disease course influenced by both major histocompatibility complex and non-major histocompatibility complex genes. Am. J. Pathol. 1996, 149: 1675-1683.
Zhao X: Dynamical Systems in Population Biology. Springer, New York; 2003.
Wang L, Wang M: Ordinary Difference Equation. Xinjiang Univ. Press, Urumqi; 1989.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant Nos. 11271312, 11001235, 10901130), the China Postdoctoral Science Foundation (Grant Nos. 20110491750, 2012T50836) and the Natural Science Foundation of Xinjiang (Grant No. 2011211B08).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
The authors declare that the study was realized in collaboration with the same responsibility. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Wang, L., Cui, Q. & Teng, Z. Global dynamics in a class of discrete-time epidemic models with disease courses. Adv Differ Equ 2013, 57 (2013). https://doi.org/10.1186/1687-1847-2013-57
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1687-1847-2013-57